
Efficacité in vitro des traitements à base d'artémisinine contre le SARS-CoV-2
Abstrait
Des traitements efficaces et abordables sont nécessaires pour les patients atteints de la maladie à coronavirus 2019 (COVID-19), causée par le coronavirus du syndrome respiratoire aigu sévère 2 (SRAS-CoV-2). Nous rapportons in vitro l'efficacité de Artemisia annua des extraits ainsi que de l'artémisinine, de l'artésunate et de l'artéméther contre le SARS-CoV-2. Ces deux derniers sont des principes actifs pharmaceutiques approuvés pour les médicaments antipaludiques.
Français La preuve de concept de l'efficacité prophylactique des extraits a été obtenue à l'aide d'un test de réduction de plaque dans des cellules VeroE6. Des études ultérieures de concentration-réponse utilisant un test antiviral à haut débit, basé sur l'immunomarquage de la glycoprotéine de pointe du SARS-CoV-2, ont révélé que le prétraitement et le traitement avec des extraits, de l'artémisinine et de l'artésunate inhibaient l'infection des cellules VeroE6 par le SARS-CoV-2. Dans les tests de traitement, l'artésunate (concentration efficace à 50 % (CE50) : 7 μg/mL) était plus puissant que les extraits de plantes testés (128-260 μg/mL) ou l'artémisinine (151 μg/mL) et l'artéméther (> 179 μg/mL), tandis que généralement la CE50 dans les tests de prétraitement était légèrement supérieure. L'indice de sélectivité (IS), calculé à partir des analyses de traitement et de viabilité cellulaire, était le plus élevé pour l'artémisinine (54), et à peu près équivalent pour les extraits (5-10), l'artésunate (6) et l'artéméther (<7). Des résultats similaires ont été obtenus sur des cellules d'hépatome humain Huh7.5. Des concentrations plasmatiques maximales d'artésunate supérieures à la CE50 peuvent être atteintes. Des études cliniques sont nécessaires pour évaluer plus en détail l'utilité de ces composés dans le traitement de la COVID-19.
Introduction
La pandémie de coronavirus du syndrome respiratoire aigu sévère 2 (SRAS-CoV-2) 1 , 2 a été associé à plus d'un million de décès dus à la maladie à coronavirus 2019 (COVID-19) dans le monde entier. 3 , 4 , 5 Cette maladie respiratoire et systémique fébrile est très contagieuse et, dans de nombreux cas, potentiellement mortelle. Le remdesivir, seul médicament antiviral dont l'efficacité a été prouvée, in vitro et son efficacité clinique, a été approuvé pour le traitement de la COVID-19. 6 Cependant, le traitement contre la COVID-19 reste largement symptomatique, et il est urgent d'identifier des antiviraux efficaces contre le SARS-CoV-2. Une approche intéressante consiste à réutiliser des médicaments déjà autorisés pour d'autres maladies. A. annua les plantes ont été utilisées pour traiter le paludisme dans la médecine traditionnelle chinoise, ainsi que dans les essais cliniques, 7 , 8 et sont largement utilisés dans de nombreux pays africains, bien que contre les recommandations de l'OMS. L'artémisinine ( Figure 1 , 1 ), une lactone sesquiterpénique avec une fraction peroxyde et l'un des nombreux composés bioactifs présents dans A. annua , est l'ingrédient actif pour traiter les infections paludiques. 9 , 10 Les dérivés de l'artémisinine artésunate ( Figure 1 , 2 ) et l'artéméther ( Figure 1 , 3 ) présentent des propriétés pharmacocinétiques améliorées et sont les principaux ingrédients pharmaceutiques actifs (IPA) des thérapies combinées antipaludiques recommandées par l’OMS, utilisées chez des millions d’adultes et d’enfants chaque année avec peu d’effets secondaires. 11 A. annua les extraits sont actifs contre différents virus, dont le SRAS-CoV. 12 , 13 , 14 Nous avons donc cherché à déterminer si A. annua les extraits, ainsi que l'artémisinine pure, l'artésunate et l'artéméther sont actifs contre le SARS-CoV-2 in vitro . Les médicaments à base d'artémisinine seraient des candidats intéressants pour le traitement de la COVID-19, compte tenu de leurs excellents profils de sécurité chez l'homme et de leur disponibilité pour une distribution mondiale à un coût relativement faible.

L'artémisinine, les dérivés API apparentés artésunate et artéméther, et l'acide chlorogénique.
Résultats
Extraits et composés
A. annua Des plantes issues d'une lignée de semences cultivées dans le Kentucky, aux États-Unis, ont été extraites à l'éthanol absolu ou à l'eau distillée à 50 °C pendant 200 minutes, comme décrit dans la section Matériels et méthodes et informations complémentaires (Figure S1). Pour une troisième préparation, afin de protéger l'artémisinine de la dégradation par les agents réducteurs, du café moulu – une source naturelle de polyphénols tels que l'acide chlorogénique ( Figure 1 , ( 4 )) qui présentent également une légère activité antivirale 15 , a été utilisé. 16 – a été coextraite avec la matière végétale à l'éthanol (voir informations complémentaires). Les solides ont été éliminés par filtration et les solvants ont été évaporés. Les matières extraites ont été dissoutes dans du diméthylsulfoxyde (DMSO) (les deux extraits à l'éthanol) ou dans un mélange DMSO:eau (3:1 pour l'extrait aqueux) puis filtrées (voir informations complémentaires pour plus de détails). L'artémisinine ( Figure 1 , ( 1 )) a été synthétisée et purifiée selon une procédure publiée, 17 tandis que l'artésunate ( Figure 1 , ( 2 )) et l'artéméther ( Figure 1 , ( 3 )) ont été obtenus à partir de sources commerciales.
Essais de réduction de plaque dans les cellules VeroE6 pour in vitro preuve de concept de l'efficacité du prétraitement A. annua extraits et artémisinine
Afin de déterminer initialement si les extraits et l'artémisinine pure étaient actifs contre le SARS-CoV-2, leur activité antivirale a été testée en prétraitant les cellules VeroE6 à différents moments pendant 120 minutes avec des concentrations sélectionnées d'extraits ou de composés avant l'infection par le premier SARS-CoV-2 européen isolé à Munich (SARS-CoV-2/humain/Allemagne/BavPat 1/2020). Le mélange virus-médicament a ensuite été retiré et les cellules ont été recouvertes d'un milieu contenant 1,3 % de carboxyméthylcellulose afin d'empêcher la libération du virus dans le milieu. Le DMSO a été utilisé comme témoin négatif. Le nombre de plages a été déterminé par immunofluorescence indirecte à l'aide d'un mélange d'anticorps dirigés contre la protéine N 18 du SARS-CoV. ou par coloration au violet de cristal. L'ajout d'éthanol ou d'eau A. annua Les extraits avant l'ajout du virus ont entraîné une réduction de la formation de plaques d'une manière dépendante de la concentration, avec des valeurs de concentration efficace médiane (CE50) estimées entre 5 et 168 μg/mL (Figure supplémentaire S2A-C). L'artémisinine a montré une faible activité antivirale avec une CE50 > 220 μg/mL (Figure supplémentaire S2D).
Efficacité de A. Annua extraits dans les antiviraux à haut débit in vitro essais sur cellules VeroE6
Des expériences de concentration-réponse utilisant l'isolat danois du SARS-CoV-2 SARS-CoV-2/human/Denmark/DK-AHH1/2020 ont été réalisées à l'aide d'un test antiviral à haut débit basé sur une plaque à 96 puits , permettant plusieurs réplications par concentration, comme décrit dans Matériels et méthodes et informations supplémentaires (Figures S3 et S4). Sept réplicats ont été mesurés à chaque concentration et une gamme de concentrations a été évaluée afin d'accroître la précision des données par rapport au test de réduction des plages, réalisé en double. Les extraits ou composés ont été ajoutés aux cellules VeroE6 respectivement 1,5 h avant (prétraitement [pt]) ou 1 h après l'infection (traitement [t]), suivis d'une incubation de deux jours du virus avec les extraits ou composés. Les deux protocoles ont donné des résultats similaires, avec des valeurs de CE50 légèrement inférieures observées pour les tests de traitement.
Les extraits éthanoliques ont montré une puissance similaire : pour A. annua seules, les CE50 étaient de 173 μg/mL (pt) et 142 μg/mL (t) et pour A. annua avec le café, les CE50 étaient de 176 μg/mL (pt) et 128 μg/mL (t) ( Figures 2 , 3 et Tableau 1 ). L'extrait aqueux était légèrement moins puissant avec une CE50 de 390 μg/mL (pt) et 260 μg/mL (t) ( Figures 2 , 3 et Tableau 1 ). Avec tous les extraits, une inhibition virale presque complète a été obtenue à des concentrations élevées : Pour le A. annua extrait éthanolique à 333 μg/mL (pt) et 444 μg/mL (t), pour le A. annua + extrait éthanolique de café à 300 μg/mL (pt) et 267 μg/mL (t), et pour le A. annua extrait aqueux à 875 μg/mL (pt) et 1009 μg/mL (t) ( Figures 2 et 3 ). Les concentrations les plus élevées évaluées utilisées dans nos tests ont été déterminées en fonction de la cytotoxicité des extraits ou des composés, car seules les concentrations entraînant une viabilité cellulaire supérieure à 90 % ont été évaluées ( Figures 2 , 3 , S5 et Tableau 1 ). Les tests de viabilité cellulaire ont révélé des concentrations cytotoxiques médianes (CC50) de 1 044 μg/mL ( A. annua extrait éthanolique), 632 μg/mL ( A. annua + extrait éthanolique de café), et 2 721 μg/mL ( A. annua extrait aqueux) ( Figures 2 , 3 , S5 et Tableau 1 ). Les indices de sélectivité (IS) ont été déterminés en divisant CC50 par CE50 et ont révélé des résultats similaires pour A. annua l'extrait éthanolique étant de 6 (pt) et 7 (t), le A. annua + extrait éthanolique de café étant 3 (pt) et 5 (t) ainsi que le A. annua extrait aqueux étant de 7 (pt) et 10 (t) ( Tableau 1 ).

Les cellules VeroE6 ensemencées la veille dans des plaques à 96 puits ont été traitées avec les concentrations spécifiées d'extraits (A) A. annua extrait éthanolique, (B) A. annua + extrait éthanolique de café, et (C) A. annua extrait aqueux ou composés artémisinine (D), artésunate (E) et artéméther (F) pendant 1,5 heure avant l'infection par le SARS-CoV-2. Après une incubation de 2 jours, les cellules infectées ont été visualisées par immunomarquage pour la glycoprotéine de pointe du SARS-CoV-2 et comptées automatiquement comme décrit dans Matériels et méthodes. Le % d'infectiosité résiduelle pour les puits individuels a été calculé en rapportant le nombre de puits traités infectés au nombre moyen de 14 puits témoins non traités infectés. Les points de données (points rouges) sont les moyennes de sept réplicats avec SEM. Les courbes dose-réponse sigmoïdales (lignes rouges) ont été ajustées et les valeurs CE50 ont été calculées dans GraphPad Prism comme décrit dans Matériels et méthodes. Le % de viabilité cellulaire et les valeurs CC50 ont été déterminés dans des essais répliqués sans infection par le SARS-CoV-2 comme décrit dans Matériels et méthodes. Les points de données (triangles bleus) sont les moyennes de 3 réplicats avec SEM. Les lignes pointillées rouges/bleues indiquent les concentrations auxquelles un effet antiviral (< 70 % d'infectiosité résiduelle) / effet cytotoxique (< 90 % de viabilité cellulaire) dû au DMSO est attendu selon la figure S6.
Les deux extraits éthanoliques ont été dilués avec du DMSO, qui a entraîné une réduction de la viabilité cellulaire inférieure à 90 % à une dilution de 1:28, mais pas à des dilutions supérieures ou égales à 1:42 (figure S6). Ainsi, la cytotoxicité observée lors de l'utilisation des extraits à des concentrations relativement élevées n'était probablement pas due au DMSO ( figures 2) . et 3 ). Le DMSO à des dilutions > 1:152, y compris celles utilisées dans les dosages antiviraux, n'a pas eu d'effet antiviral, défini comme une réduction de l'infectiosité résiduelle à < 70 % (Figure S6). Par conséquent, l'effet antiviral observé des extraits testés n'était probablement pas dû au DMSO. Un extrait de café pur, dont on estime qu'il contient des concentrations de café 2,5 fois supérieures à celles de l'extrait de café pur. A. annua + l'extrait éthanolique de café n'a pas entraîné de réduction de la viabilité cellulaire à < 90 % à des dilutions ≥ 1:28 (Figure S7). La cytotoxicité observée lors de l'utilisation de A. annua + l'extrait de café à des concentrations relativement élevées n'était probablement pas causé par le café ( Figures 2 et 3 ). Il est intéressant de noter que l'extrait de café seul a montré une certaine activité antivirale à des dilutions ≤ 1:273 (Figure S7). Ainsi, l'effet antiviral observé de A. annua + l'extrait de café peut être influencé par le café.

Les cellules VeroE6 ensemencées la veille dans des plaques à 96 puits ont été infectées par le SARS-CoV-2 et après 1 heure d'incubation traitées avec les concentrations spécifiées d'extraits (A) A. annua extrait éthanolique, (B) A. annua + extrait éthanolique de café, et (C) A. annua Extrait aqueux ou composés d'artémisinine (D), d'artésunate (E) et d'artéméther (F). Après une incubation de 2 jours, les cellules infectées ont été visualisées par immunomarquage pour la glycoprotéine de pointe du SARS-CoV-2 et comptées automatiquement comme décrit dans Matériels et méthodes. Le % d'infectiosité résiduelle pour chaque puits a été calculé en rapportant le nombre de puits traités infectés au nombre moyen de 14 puits témoins infectés non traités. Les points de données (points rouges) sont les moyennes de sept réplicats avec SEM. Les courbes dose-réponse sigmoïdales (lignes rouges) ont été ajustées et les valeurs CE50 ont été calculées dans GraphPad Prism comme décrit dans Matériels et méthodes. Le % de viabilité cellulaire et les valeurs CC50 ont été déterminés dans des essais répliqués sans infection par le SARS-CoV-2 comme décrit dans Matériels et méthodes. Les points de données (triangles bleus) sont les moyennes de trois réplicats avec SEM. Les lignes pointillées rouges/bleues indiquent les concentrations auxquelles un effet antiviral (< 70 % d'infectiosité résiduelle) / effet cytotoxique (< 90 % de viabilité cellulaire) dû au DMSO est attendu selon la figure S6.
Efficacité de l'artémisinine et de ses dérivés dans les antiviraux à haut débit in vitro essais sur cellules VeroE6
A. annua Les plantes contiennent, en plus de nombreux autres composés bioactifs, de l'artémisinine, responsable des puissantes activités antipaludiques de A. annua . Afin de déterminer si l'artémisinine est le composant actif responsable des activités antivirales des extraits végétaux décrits ci-dessus, le composé pur et ses dérivés synthétiques ont été testés lors d'essais de prétraitement et de traitement. L'artémisinine s'est révélée active dans les essais de dépistage du SARS-CoV-2 avec des CE50 de 238 μg/mL (pt) et 151 μg/mL (t) ( Figures 2 , 3 , et Tableau 1 ). Une inhibition virale quasi complète a été obtenue dans les deux essais à la concentration la plus élevée évaluée, soit 893 (pt) et 1 208 μg/mL (t). L'indice de réactivité (IS) de l'artémisinine est relativement élevé, soit 34 (pt) et 54 (t), sur la base d'une CC50 de 8 216 μg/mL ( Figures 2 , 3 , S6 et Tableau 1 ). La cytotoxicité observée de l'artémisinine semble être au moins partiellement causée par le DMSO, car la cytotoxicité n'a été observée qu'à des dilutions de médicament où le DMSO s'est avéré réduire la viabilité cellulaire ( Figures 2 , 3 et S6). Les effets antiviraux observés lors de l'utilisation d'artémisinine à des concentrations relativement élevées n'étaient probablement pas dus au diluant DMSO ( Figures 2 , 3 et S6).
L'artésunate, dérivé synthétique de l'artémisinine, l'IPA des thérapies antipaludiques de première intention recommandées par l'OMS avec des propriétés pharmacocinétiques améliorées, a montré la plus grande puissance de tous les composés testés, avec une CE50 de 12 μg/mL (pt) et 7 μg/mL (t) ( Figures 2 et 3 ). Lors de l'essai de traitement, une inhibition virale quasi complète a été obtenue à la concentration la plus élevée évaluée (15 μg/mL), d'après les données de cytotoxicité, contre 69 % d'inhibition à cette concentration lors de l'essai de prétraitement. Des concentrations plus élevées d'artésunate n'ont pas été utilisées compte tenu de sa cytotoxicité dans cet essai (CC50 : 41 μg/mL) ( Figures 2 , 3 , S5 et Tableau 1 ). Des SI de 3 (pt) et 6 (t) ont été calculés ( Tableau 1 ). La cytotoxicité et les effets antiviraux observés lors de l'utilisation d'artésunate à des concentrations relativement élevées n'étaient probablement pas dus au diluant DMSO ( Figures 2 , 3 et S6).
L'artéméther, un autre dérivé de l'artémisinine utilisé dans le monde entier comme ingrédient actif dans les médicaments contre le paludisme, n'a pas montré d'effet antiviral significatif à des concentrations allant jusqu'à 179 μg/mL ( Figures 2 et 3 ). Compte tenu de la cytotoxicité de l'artéméther (CC50 de 1 220 μg/mL), un SI < 7 a été calculé ( Figures 2 , 3 , S5 et Tableau 1 ). La cytotoxicité observée lors de l'utilisation d'artéméther à des concentrations relativement élevées n'était probablement pas due au diluant DMSO ( Figures 2 , 3 et S6).
Efficacité du traitement à base d'artémisinine dans les antiviraux à haut débit in vitro dosages utilisant des cellules Huh7.5
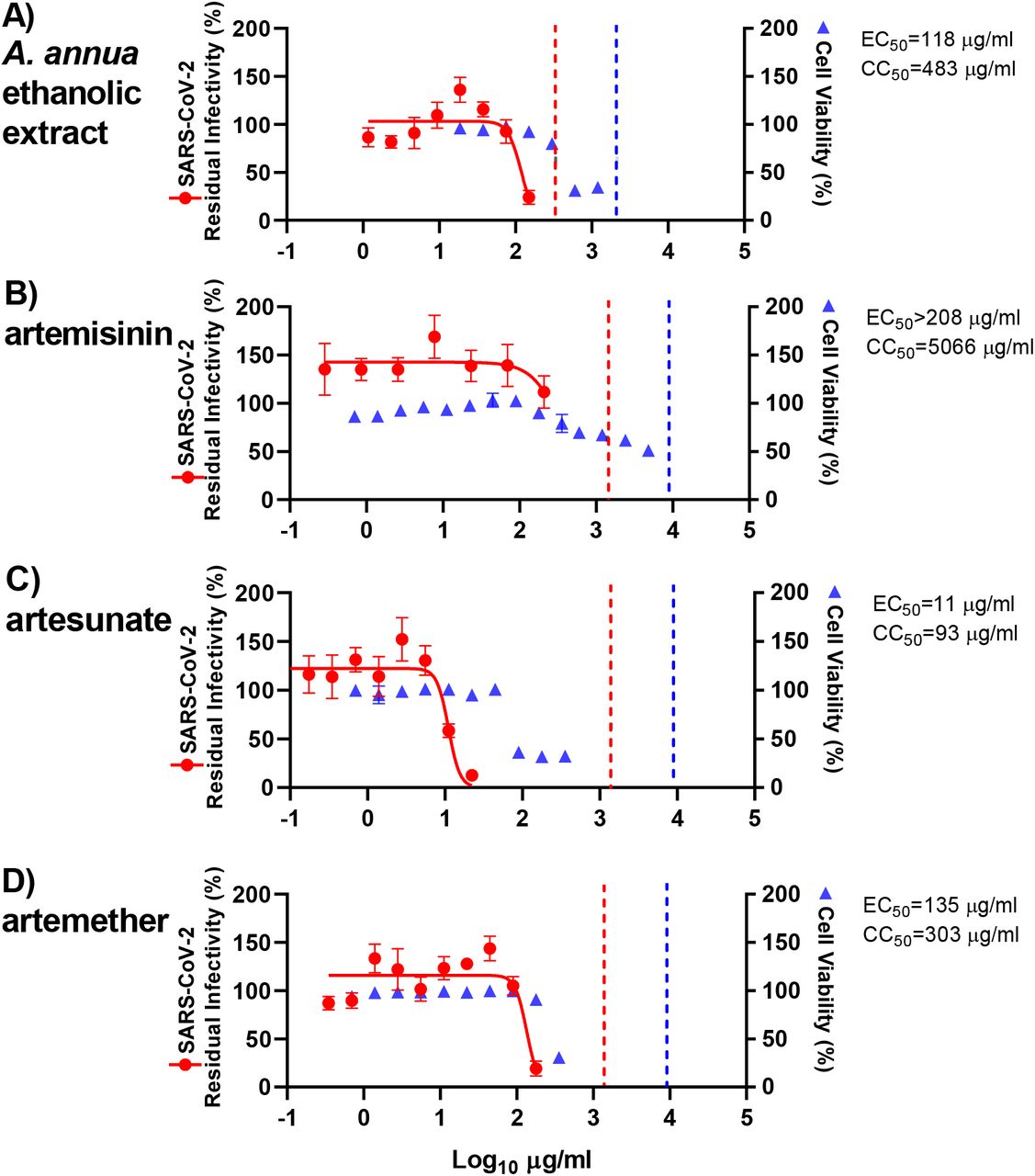
L'activité antivirale observée lors de ces tests est influencée par la capacité des composés purs et des composés contenus dans les extraits à pénétrer dans les cellules, ainsi que par leur métabolisme. Afin d'exclure toute différence majeure d'activité des extraits et des composés dans les cellules humaines, des tests de traitement ont également été réalisés sur des cellules d'hépatome humain Huh7.5, en ajoutant des extraits ou des composés aux cellules immédiatement après l'infection. Globalement, l'éthanol a été utilisé. A. annua L'extrait, l'artémisinine, l'artésunate et l'artéméther ont montré une efficacité similaire dans les cellules Huh7.5 par rapport aux cellules VeroE6. L'artésunate (EC50 : 11 µg/mL) s'est à nouveau révélé être le composé le plus puissant, avec une inhibition virale quasi complète à 22 µg/mL et un SI de 8, déterminé par une CC50 de 93 µg/mL ( Figures 4 , S8 et Tableau 1 ). L'artéméther (CE50 : 135 μg/mL) avec une inhibition virale presque complète à 179 μg/mL, avait un SI de seulement 2, basé sur une CC50 de 303 μg/mL ( Figures 4 , S8 et Tableau 1 ). Dans les cellules Huh7.5, la CE50 pour l'éthanol A. annua l'extrait était de 118 μg/mL, avec une inhibition virale de 76 % à la concentration la plus élevée évaluée (150 μg/mL), comme déterminé par les données de cytotoxicité ; le CC50 était de 483 μg/mL et le SI était de 4 ( Figures 4 , S8 et Tableau 1 ). L'artémisinine n'a montré aucune inhibition virale significative à la concentration évaluée la plus élevée (208 μg/mL) et un SI < 24, basé sur une CC50 de 5 066 μg/mL ( Figures 4 , S8 et Tableau 1 ).

Les cellules Huh7.5 ensemencées la veille dans des plaques à 96 puits ont été infectées par le SARS-CoV-2 et traitées directement avec les concentrations spécifiées d'extrait (A) A. annua extrait éthanolique ou composés artémisinine (D), artésunate (E) et artéméther (F). Après une incubation de 3 jours, les cellules infectées ont été visualisées par immunomarquage pour la glycoprotéine de pointe du SARS-CoV-2 et comptées automatiquement comme décrit dans Matériels et méthodes. Le % d'infectiosité résiduelle pour les puits individuels a été calculé en rapportant le nombre de puits traités infectés au nombre moyen de 14 puits témoins infectés non traités. Les points de données (points rouges) sont les moyennes de sept réplicats avec SEM. Les courbes dose-réponse sigmoïdales (lignes rouges) ont été ajustées et les valeurs CE50 ont été calculées dans GraphPad Prism comme décrit dans Matériels et méthodes. Le % de viabilité cellulaire et les valeurs CC50 ont été déterminés dans des essais répliqués sans infection par le SARS-CoV-2 comme décrit dans Matériels et méthodes. Les points de données (triangles bleus) sont les moyennes de 3 réplicats avec SEM. Les lignes pointillées rouges/bleues indiquent les concentrations auxquelles un effet antiviral (< 70 % d'infectiosité résiduelle) / effet cytotoxique (< 90 % de viabilité cellulaire) dû au DMSO est attendu selon la figure S9.
Dans les cellules Huh7.5, le DMSO a entraîné une réduction de la viabilité cellulaire inférieure à 90 % à une dilution de 1:28, mais pas à des dilutions ≥ 1:56 (Figure S9). Ainsi, la cytotoxicité observée lors de l'utilisation de l'extrait éthanolique ou des composés purs à des concentrations relativement élevées n'était probablement pas due au DMSO ( Figure 4 ). Le DMSO à des dilutions > 1:179, y compris celles utilisées dans les dosages antiviraux, n'a eu aucun effet antiviral (Figure S9). Par conséquent, l'effet antiviral observé de l'extrait éthanolique A. annua l'extrait et les composés purs n'ont probablement pas été causés par le DMSO.
Discussion
Ici, nous démontrons la in vitro l'efficacité des traitements à base d'artémisinine contre le SRAS-CoV-2. Initialement, plusieurs A. annua Des extraits de SARS-CoV-2, ainsi que l'artémisinine, ont été examinés pour leur activité antivirale à l'aide d'un test de réduction de plaques dans un contexte de prétraitement utilisant une souche allemande de SARS-CoV-2 de Munich. Sur la base de ces résultats, trois A. annua Des extraits et de l'artémisinine, de l'artésunate et de l'artéméther synthétiques purs ont été étudiés en détail pour établir des courbes concentration-réponse pour les extraits et les composés destinés aux contextes de prétraitement et de traitement utilisant une souche danoise de SARS-CoV-2 de Copenhague.
Les dosages antiviraux à haut débit ont facilité le test des concentrations de médicament en plusieurs réplicats, ce qui a permis d'obtenir des valeurs de CE50 précises. Les valeurs de CE50 obtenues avant traitement étaient légèrement supérieures à celles obtenues après traitement, probablement en raison d'un impact négatif de la préincubation sur la stabilité des extraits et des composés purs. En général, les valeurs de CE50 dépendent du dosage utilisé. Bien que le type de dosage utilisé, avec un traitement unique et l'incubation ultérieure du virus et du médicament, soit à la pointe de la technologie pour les mesures d'efficacité antivirale, des modifications du dosage, telles que l'administration répétée du traitement, pourraient entraîner des valeurs de CE50 légèrement différentes. Comme la substance antivirale active pourrait être un métabolite de l'artémisinine, les dérivés et extraits d'artémisinine pouvant être considérés comme des promédicaments, nous avons utilisé la lignée cellulaire humaine Huh7.5 pour confirmer la CE50 déterminée sur les cellules VeroE6.
Alors que A. annua Les extraits ont été considérés comme des « thérapies combinées naturelles » car ils contiennent plusieurs composés bioactifs, 21 L'OMS déconseille l'utilisation de formes non pharmaceutiques d'artémisinine comme option thérapeutique contre le paludisme en raison du manque de normalisation de son approvisionnement et de sa préparation, ce qui implique des risques d'efficacité sous-optimale et de développement de résistance. 22 Dans ce contexte, il est important de noter que les extraits utilisés dans cette étude ont été préparés à partir de plantes cultivées dans des conditions optimisées et standardisées, de manière à ce que les concentrations du matériel extrait soient reproductibles.
Il est intéressant de noter que nous avons constaté que les extraits de café présentaient in vitro Efficacité contre le SARS-CoV-2. Bien que des études de modélisation aient suggéré que des ingrédients du café, tels que l'acide chlorogénique, l'acide caféique et les tanins, présentent une activité contre le SARS-CoV-2, 23 nous fournissons ici in vitro Il n'existe aucune preuve d'un tel effet. Des études ultérieures sont nécessaires pour élucider en détail l'effet du café et de ses principes actifs sur le SARS-CoV-2. De plus, de futures études pourraient déterminer si A. annua ou les ingrédients actifs et les extraits de café présentent des effets antagonistes, additifs ou synergiques sur le SARS-CoV-2.
Lors de la compilation des données pour cette étude, Cao et al. ont rapporté l’efficacité des dérivés de l’artémisinine contre un isolat de SARS-CoV-2 de Wuhan dans les cellules VeroE6.24 Bien que les extraits n'aient pas été étudiés, l'efficacité de l'artésunate était similaire, avec une CE50 de 13 μM contre 18 μM dans notre essai de traitement VeroE6. Il est intéressant de noter que les CE50 de l'artéméther et de l'artémisinine étaient 8 fois et plus de 8 fois supérieures dans notre essai de traitement VeroE6 par rapport aux valeurs rapportées par Cao. et al . Cette différence pourrait être due à la nature du test utilisé. Le test utilisé par Cao Français et al . était basé sur des déterminations d'ARN viral, l'inoculum viral a été retiré après l'infection / avant le traitement et peut-être plus important encore, le test a été terminé 24 heures après l'infection, ce qui devrait entraîner une CE50 comparativement plus faible pour les composés ayant un effet antiviral immédiat mais une capacité limitée à contrôler le virus, soit en raison d'une efficacité antivirale limitée, soit en raison d'une stabilité limitée pendant une période de traitement de 48 heures. Il est important de noter que nous rapportons pour la première fois l'efficacité des composés dans une lignée cellulaire humaine (Huh7.5) en plus de l'efficacité dans la lignée cellulaire de singe VeroE6. Enfin, dans notre étude, nous confirmons l'efficacité du traitement à base d'artémisinine pour deux souches européennes de SARS-CoV-2 provenant d'Allemagne et du Danemark, qui sont plus étroitement liées à la majorité des souches de SARS-CoV-2 circulant dans le monde que la souche de Wuhan.
L'artésunate, l'ingrédient actif des traitements antipaludiques approuvés par la Food and Drug Administration (FDA), a montré la plus grande efficacité contre le SARS-CoV-2 parmi les extraits et composés purs testés sur cellules VeroE6 et Huh7.5. Les extraits se sont révélés plus puissants que l'artémisinine et l'artéméther, dont l'efficacité antivirale était limitée compte tenu des résultats obtenus sur cellules VeroE6 et Huh7.5 . Avec un indice de sécurité (IS) inférieur à 10, à l'exception de l'artémisinine, la marge thérapeutique est relativement faible. Il convient de noter que certains médicaments, comme la digoxine, dont l'IS peut descendre jusqu'à 2, sont utilisés avec succès en clinique. Parmi les extraits et composés purs testés, seul l'artésunate a montré des valeurs de CE50 dans la fourchette des concentrations plasmatiques et tissulaires cliniquement réalisables . Après administration intraveineuse des doses usuelles de 2 à 2,4 mg/kg, les concentrations plasmatiques maximales (Cmax) observées chez les patients se situaient entre 19,4 et 29,7 μg/mL. Sur la base de ces observations et de nos données de traitement sur cellules VeroE6 et Huh7.5, les valeurs Cmax/CE50 calculées se situent entre 2,5 et 4,2. Dans les études animales suivant l'administration d'une dose unique d'artésunate, les concentrations tissulaires, notamment pulmonaires, rénales, intestinales et spléniques, étaient plusieurs fois supérieures aux concentrations plasmatiques. En revanche, après administration d’artémisinine, d’artéméther et A. annua Dans les thés, des valeurs de Cmax comprises entre 311 et 776 ng/mL ont été rapportées, ce qui est près de trois ordres de grandeur inférieur aux valeurs de CE50 pour le SARS-CoV-2. Les concentrations plasmatiques et tissulaires peuvent être atteintes avec des concentrations standardisées. A. annua les extraits à haute teneur en artémisinine utilisés dans cette étude doivent encore être déterminés. In vivo , des effets immunomodulateurs des traitements à base d’artémisinine ont été rapportés pour cette classe de médicaments.28 De tels effets qui peuvent impliquer la signalisation des cytokines ne peuvent pas être surveillés in vitro des analyses ont été réalisées ici et devront être soigneusement étudiées lors d'évaluations cliniques ultérieures.
Matériels et méthodes
Extraction
Le solvant (250 ml d'éthanol ou d'eau distillée) a été chauffé à 50 °C dans un erlenmeyer. Des matières végétales séchées (50 g pour l'éthanol, 25 g pour l'eau) ou des matières végétales séchées et du café prémoulu (50 g, 50 g) ont été ajoutés au solvant et agités pendant 200 minutes. Le mélange a été filtré et les matières solides lavées à l'éthanol frais ou à l'eau. Le solvant a été éliminé par évaporation rotative et les matières solides ont été conservées à −30 °C avant la préparation de l'échantillon.
Préparation des échantillons
L'extrait sec a été réchauffé à température ambiante. La masse d'échantillon requise a été prélevée à l'aide d'une spatule. Du DMSO (3 mL, extraits à l'éthanol) ou du DMSO:eau (3:1, 8 mL d'extrait aqueux) a été ajouté et le mélange a été chauffé (40 °C) pour assurer la solvatation. La solution a été filtrée à l'aide d'un filtre seringue et conservée dans un flacon à fermeture rapide.
Culture cellulaire
À la FU Berlin, les cellules rénales VeroE6 de singe vert africain (ATCC CRL-1586) ont été maintenues à 37 °C avec 5 % de CO 2 dans un milieu essentiel minimum (MEM ; PAN Biotech, Aidenbach, Allemagne) supplémenté avec 10 % de sérum fœtal bovin (PAN Biotech), 100 UI/mL de pénicilline G et 100 μg/mL de streptomycine (Carl Roth, Karlsruhe, Allemagne).
Au CO-HEP, cellules VeroE6 de rein de singe vert africain (don de J. Dubuisson) ainsi que cellules d'hépatome humain Huh7.5 28 ont été maintenus à 37 °C avec 5 % de CO 2 dans du milieu modifié de Dulbecco (DMEM) (Invitrogen, Paisley, Royaume-Uni) contenant 10 % de sérum fœtal bovin (SVF) inactivé par la chaleur (Sigma, Saint Louis, Missouri, États-Unis) et 100 U/mL de pénicilline + 100 μL de streptomycine (Gibco/Invitrogen Corporation, Carlsbad, Californie, États-Unis). Les cellules ont été repiquées tous les 2 à 3 jours à l'aide de trypsine (Sigma, Saint Louis, Missouri, États-Unis) afin de maintenir une couche cellulaire sous-confluente.
Isolats de virus
L'isolat SARS-CoV-2 BavPat 1 (SARS-CoV-2/humain/Allemagne/BavPat 1/ 2020 a été fourni par le Dr Daniela Niemeyer et le Dr Christian Drosten (Charité, Berlin, Allemagne) et obtenu lors d'une épidémie à Munich, en Allemagne, en février 2020 (BetaCoV/Allemagne/BavPat1/2020).
Le virus SARS-CoV-2/humain/Danemark/DK-AHH1/2020 pour les études de culture cellulaire a été obtenu après inoculation de cellules VeroE6 avec un échantillon d'écouvillon de patient, propagation du virus dans les cellules VeroE6 et génération d'un stock de 2e passage viral confirmé par séquence avec un titre d'infectiosité de 5,5 log TCID50/mL comme décrit dans Ramirez et al . 20
Test antiviral de réduction de plaque
L'activité antivirale des dérivés de l'artémisinine a été évaluée sur des cellules VeroE6 cultivées pendant la nuit dans des plaques à 12 puits (Sarstedt) à une densité d'environ 5 × 10 5 cellules/puits. Les cellules ont été incubées en présence de dilutions en série au dixième des composés pendant 15, 30, 60 ou 120 minutes, avant l'ajout du virus à une concentration d'environ 200 unités formant des plages (UFP) par puits pendant 120 minutes. Le mélange virus-médicament a été retiré et les cellules ont été recouvertes de MEM-FBS contenant 1,3 % de carboxyméthylcellulose afin d'empêcher la libération du virus dans le milieu. Du DMSO dans du milieu de culture cellulaire à une dilution de 1:100 (la concentration la plus élevée par rapport aux préparations d'extraits/composés) a été utilisé comme témoin négatif, et le nombre de plages virales a été déterminé par comptage manuel des plages après immunofluorescence indirecte (IF) à l'aide d'un mélange d'anticorps dirigés contre la protéine N 18 du SARS-CoV. ou après coloration au violet de cristal. Pour l'IF, les cellules ont été fixées avec du formol à 4 % et perméabilisées avec du Triton X-100 à 0,25 %. La liaison non spécifique a été bloquée avec du FBS à 1 % dans du tampon phosphate salin (PBS) contenant 0,25 % de Triton X-100 (PBS-T) à température ambiante pendant 30 minutes. Les cellules ont été incubées avec les anticorps monoclonaux anti-N (dilution 1:25 dans du PBS-T) pendant 45 minutes, puis incubées avec un anticorps secondaire (anticorps de chèvre anti-souris marqué à l'Alexa 488 à une dilution 1:500 ; Thermo Fisher). Dans chaque test, chaque concentration a été testée dans une culture répliquée ; 5 cultures infectées et témoins traitées au DMSO ont été incluses dans chaque test. Le nombre de plaques enregistré dans chaque culture infectée traitée a été rapporté au nombre moyen des cinq cultures témoins pour calculer le nombre de plaques en pourcentage par rapport au témoin. Deux tests indépendants ont été réalisés. Les points de données correspondent aux moyennes de deux cultures répliquées issues de deux tests indépendants, les barres d'erreur reflétant les écarts types (ET) (figure S2). Les concentrations sélectionnées n'ont été testées que dans un seul des tests et, pour ces points de données, sont basées sur des réplicats uniques. Le MOI pour l'infection a été choisi avec un objectif moyen de 150 à 250 plaques par culture.
Essai antiviral de prétraitement et de traitement à haut débit dans les cellules VeroE6
Des tests antiviraux à 96 puits dans des cellules VeroE6 ont été développés sur la base de tests précédemment établis pour l'évaluation de l'efficacité des antiviraux contre le virus de l'hépatite C. 30 , 31 Les cellules VeroE6 ont été ensemencées à raison de 10 000 cellules par puits dans des plaques 96 puits recouvertes de poly-D-lysine (Thermo Fisher Scientific, Rochester, NY, États-Unis). Pour les tests de prétraitement, le lendemain, le milieu a été remplacé par un milieu contenant des extraits ou des composés, en ajoutant 50 µL par puits. Après 1,5 h d'incubation à 37 °C et 5 % de CO₂ , les cellules ont été inoculées avec le virus SARS-CoV-2/humain/Danemark/DK-AHH1/2020 à une MOI de 0,0016 en ajoutant 50 µL de stock viral dilué par puits, ce qui a permis d'obtenir les concentrations spécifiées d'extraits ou de composés. Pour les tests de traitement, le lendemain, le milieu a été remplacé par 50 µL de milieu frais par puits. Français Ensuite, les cellules ont été inoculées avec SARS-CoV-2/humain/Danemark/DK-AHH1/2020 à MOI 0,0016 en ajoutant 50 μL de stock de virus dilué par puits. Après 1 heure d'incubation à 37 °C avec 5 % de CO 2 , 50 μL de milieu contenant des extraits ou des composés ont été ajoutés, ce qui a donné les concentrations spécifiées ; alternativement, 50 μL de milieu contenant un diluant (DMSO) ou un additif (extrait de café) ont été ajoutés, ce qui a donné les dilutions spécifiées. Pour les deux tests, chaque concentration/dilution a été testée en sept réplicats ; 14 puits infectés et non traités ainsi que 12 puits témoins non infectés et non traités ont été inclus dans chaque test. Après 48 ± 2 heures d'incubation à 37 °C et 5 % de CO 2 , les cultures ont été immunocolorées pour la glycoprotéine de pointe du SARS-CoV-2 et évaluées comme décrit ci-dessous.
Essai antiviral de traitement à haut débit dans les cellules Huh7.5
Les cellules Huh7.5 ont été ensemencées à raison de 8 000 cellules par puits dans des plaques à fond plat de 96 puits (Thermo Fisher Scientific, Roskilde, Danemark). Le lendemain, les cellules ont été inoculées avec le virus SARS-CoV-2/humain/Danemark/DK-AHH1/2020 à une MOI de 0,0198 en ajoutant 50 μL de stock viral dilué par puits. Immédiatement après, 50 μL de milieu contenant des extraits ou des composés ont été ajoutés, ce qui a permis d'obtenir les concentrations spécifiées ; alternativement, 50 μL de milieu contenant un diluant (DMSO) ont été ajoutés, ce qui a permis d'obtenir les dilutions spécifiées. Chaque concentration a été testée en sept réplicats ; 14 puits infectés et non traités ainsi que 12 puits témoins non infectés et non traités ont été inclus dans le test. Après 72 ± 2 heures d'incubation à 37 °C et 5 % de CO 2 , les cultures ont été immunocolorées pour la glycoprotéine de pointe du SARS-CoV-2 et évaluées comme décrit ci-dessous.
Immunocoloration et évaluation de plaques à 96 puits pour des dosages antiviraux à haut débit
Français Les cellules ont été fixées et le virus a été inactivé par immersion des plaques dans du méthanol (JTBaker, Gliwice, Pologne) pendant 20 min. Sauf indication contraire, l'immunomarquage a été réalisé à température ambiante. Les plaques ont été lavées deux fois avec du PBS (Sigma, Gillingham, Royaume-Uni) contenant 0,1 % de Tween-20 (Sigma, Saint Louis, Missouri, États-Unis). L'activité de la peroxydase endogène a été bloquée par incubation avec 3 % de H2O2 pendant dix minutes, suivie de deux lavages avec du PBS contenant 0,1 % de Tween-20 et d'un blocage avec du PBS contenant 1 % d'albumine sérique bovine (Roche, Mannheim, Allemagne) et 0,2 % de poudre de lait écrémé (Easis, Aarhus, Danemark) pendant 30 minutes. Français Après le retrait de la solution de blocage, les plaques ont été incubées avec l'anticorps monoclonal chimérique de pointe SARS-CoV-2 (Sino Biological #40150-D004, Pékin, Chine) dilué à 1:5000 dans du PBS contenant 1 % d'albumine sérique bovine et 0,2 % de lait écrémé en poudre pendant la nuit à 4 °C. Après deux lavages avec du PBS contenant 0,1 % de Tween-20, les plaques ont été incubées avec l'anticorps secondaire F(ab')2-anticorps secondaire anti-IgG humaine de chèvre à adsorption croisée Fc, HRP (Invitrogen #A24476, Carlsbad, CA, États-Unis) ou l'anticorps F(ab')2 anti-IgG humaine de chèvre - Fc (HRP), pré-adsorbé (Abcamab#98595, Cambridge, Royaume-Uni) dilué à 1:2000 dans du PBS contenant 1 % d'albumine sérique bovine et 0,2 % de lait écrémé en poudre pendant 2 h. Après deux lavages avec du PBS contenant 0,1 % de Tween-20, la glycoprotéine de spicule du SARS-CoV-2 a été visualisée à l'aide du substrat DAB (Immunologic n° BS04-110, Duiven, Pays-Bas). Les cellules positives à la protéine de spicule ont été comptées automatiquement à l'aide d'un analyseur UV ImmunoSpot série 5 (CTL Europe GmbH, Bonn, Allemagne), comme décrit. 30 , 31 , 32 Français Le nombre moyen de 12 puits témoins non infectés et non traités, qui était généralement < 50, a été soustrait du nombre de chaque puits infecté. Les nombres enregistrés dans chaque puits traité infecté ont été rapportés au nombre moyen de 14 puits témoins non traités infectés pour calculer le % d'infectiosité résiduelle. Les points de données sont les moyennes de sept réplicats avec des erreurs standard des moyennes (SEM). Des courbes dose-réponse sigmoïdales ont été ajustées et les valeurs CE50 ont été calculées avec GraphPad Prism 8.0.0 en utilisant une contrainte inférieure de 0 et la formule Y = Haut/(1+10^((LogEC50-X)*Pente)). Le MOI pour l'infection a été choisi en visant une moyenne de 3000 à 4000 comptes par puits pour les cellules VeroE6 et une moyenne de 300 à 600 comptes par puits pour les cellules Huh7.5 moins permissives dans les puits témoins infectés et non traités à la fin des tests respectifs. Des images représentatives de 96 puits provenant d'essais dans des cellules VeroE6 sont présentées dans la figure S3 et des images représentatives de puits individuels sont présentées dans la figure S4.
Essais de viabilité cellulaire dans les cellules VeroE6 et Huh7.5
Pour évaluer les effets cytotoxiques des extraits, composés, diluants (DMSO) et additifs (extrait de café) testés, la viabilité cellulaire a été surveillée à l'aide du CellTiter 96 ® Essai de prolifération cellulaire en solution aqueuse (Promega, Madison, Wisconsin, États-Unis). Des cellules VeroE6 ou Huh7.5 ont été ensemencées à raison de 10 000 ou 8 000 cellules par puits dans des plaques 96 puits à fond plat, respectivement (Thermo Fisher Scientific, Roskilde, Danemark). Le lendemain, le milieu a été renouvelé pour contenir les concentrations spécifiées d'extraits, de composés ou de dilutions de DMSO et d'extrait de café, en ajoutant 100 μL par puits. Chaque concentration ou dilution a été testée en 3 réplicats ; au moins 6 puits témoins non traités ont été inclus dans l'essai. Pour les cellules VeroE6, après 48 ± 2 h d'incubation à 37 °C et 5 % de CO₂, et pour les cellules Huh7.5, après 72 ± 2 h d'incubation à 5 % de CO₂ , 20 μL de CellTiter 96 ® ont été ajoutés. Un réactif Aqueous One Solution a été ajouté à chaque puits et les plaques ont été incubées pendant 1,5 à 2 h à 37 °C et 5 % de CO₂ , avant l'enregistrement de l'absorbance à 492 nm à l'aide d'un lecteur de plaques FLUOstar OPTIMA 96 puits (BMG LABTECH, Offenburg, Allemagne). L'absorbance enregistrée dans chaque puits a été rapportée à l'absorbance moyenne des puits témoins non traités pour calculer le pourcentage de viabilité cellulaire. Les points de données sont des moyennes de triplicats avec SEM. Des courbes dose-réponse sigmoïdales ont été ajustées et les valeurs de concentration cytotoxique médiane (CC50) ont été calculées avec GraphPad Prism 8.0.0 en utilisant une contrainte inférieure de 0 et la formule Y = Haut/(1+10^((LogCC50-X)*Pente)) comme spécifié plus en détail dans les figures S5 et S8. Pour exclure les effets cytotoxiques aux concentrations sélectionnées en fonction des tests de viabilité cellulaire en présence d’une infection virale, les puits de culture dans les tests antiviraux ont été inspectés manuellement au microscope optique.
Contributions des auteurs
KG et PHS ont conçu ce projet. KG, YZ, SR, JB, KO, JMG et PHS ont conçu les expériences. KG a effectué les extractions et les préparations d'échantillons. KO et JT ont réalisé les tests de réduction des plages. YZ, SR, LP, UF, SF et AO ont contribué à l'isolement du SARS-CoV-2/humain/Danemark/DK-AHH1/2020 et ont établi des systèmes expérimentaux. YZ a réalisé les tests antiviraux à haut débit. Tous les auteurs ont analysé les données. Tous les auteurs ont contribué et discuté du manuscrit.
Informations sur l'auteur
Toute correspondance relative aux tests de réduction de plaques doit être adressée à KO ( no34{at}cornell.edu ). Toute correspondance relative aux tests antiviraux à haut débit et aux tests de viabilité cellulaire doit être adressée à JMG ( jgottwein{at}sund.ku.dk ). Toute correspondance concernant les extractions et les composés purs doit être adressée à PHS ( peter.seeberger{at}mpikg.mpg.de ).
Conflit d'intérêts
KG est le directeur d'ArtemiLife, Inc. KG et PHS détiennent une participation financière importante dans ArtemiFlow GmbH, qui est actionnaire d'ArtemiLife, Inc.
Remerciements
Nous remercions la Société Max Planck pour son soutien financier. Ce travail a bénéficié d'une bourse de doctorat du Conseil chinois des bourses (YZ) et d'une subvention de l'Agence danoise pour la science et l'enseignement supérieur (JB). Nous remercions le Dr Bjarne Ø. Lindhardt (hôpital universitaire de Copenhague, Hvidovre) et le professeur Carsten Geisler (université de Copenhague) pour leur soutien de l'hôpital de Hvidovre et de l'université de Copenhague. Nous remercions Lotte Mikkelsen, Anna-Louise Sørensen et Pia Pedersen (hôpital universitaire de Copenhague, Hvidovre) pour leur aide en laboratoire. Nous remercions le Dr Christoph Rademacher et Felix Fuchsberger pour leurs précieux échanges. Nous remercions également ArtemiLife Inc. pour la mise à disposition des informations. A. annua matériel végétal. Nous remercions le Pr Jean Dubuisson et le Dr Sandrine Belouzard pour la mise à disposition des cellules VeroE6.
- Liste des abréviations
- (API)
- Ingrédients pharmaceutiques actifs
- (CC50)
- concentration cytotoxique médiane
- (COVID 19)
- maladie du coronavirus 2019
- (DMSO)
- diméthylsulfoxyde
- (EC50)
- concentration efficace médiane
- (FBS)
- sérum fœtal bovin
- (FDA)
- Administration des aliments et des médicaments
- (SI)
- immunofluorescence
- (PBS)
- solution saline tamponnée au phosphate
- (SD)
- écart type
- (SEM)
- erreur standard de la moyenne
- (SI)
- indice de sélectivité
- (SRAS-CoV-2)
- coronavirus 2 du syndrome respiratoire aigu sévère

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}